Современные проблемы науки и образования. Транспорт углекислоты кровью
Из венозной крови можно извлечь 55-58 об.% углекислого газа. Большая часть СО2, извлекаемого из крови, происходит из имеющихся в плазме и эритроцитах солей угольной кислоты и только около 2,5 об.% углекислого газа растворено и около 4-5об.% находится в соединении с гемоглобином в виде карбогемоглобина.
Образованно угольной кислоты из углекислого газа происходит в эритроцитах, где содержится фермент карбоангидраза, являющийся мощным катализатором, ускоряющим реакцию гидратации СО2.
Связывание углекислого газа кровью в капиллярах большого круга. Углекислый газ, образующийся в тканях, диффундирует в кровь кровеносных капилляров, так как напряжение СО2 в тканях значительно превышает его напряжение в артериальной крови. Растворяющийся в плазме СО2 диффундирует внутрь эритроцита, где под влиянием карбоангидразы он мгновенно превращается в угольную кислоту,
Согласно расчетам, активность карбоангидразы в эритроцитах такова, что реакция гидратации углекислоты ускоряется в 1500-2000 раз. Так как весь углекислый газ внутри эритроцита превращается в угольную кислоту, то напряжение СО2 внутри эритроцита близко к нулю, поэтому все новые и новые количества СО2 поступают внутрь эритроцита. В связи с образованием угольной кислоты из СО3 в эритроците концентрация ионов НСО3" возрастает, и они начинают диффундировать в плазму. Это возможно потому, что поверхностная мембрана эритроцита проницаема для анионов. Для катионов мембрана эритроцита практически непроницаема. Взамен ионов НСО3" в эритроциты входит ион хлора. Переход ионов хлора из плазмы внутрь эритроцита освобождает в плазме ионы натрия, которые связывают поступающие нз эритроцита ионы НСО3, образуя NaHCО3 Химический анализ плазмы венозной крови показывает значительное увеличение в ней бикарбоната.
Накопление внутри эритроцита анионов приводит к повышению осмотического давления внутри эритроцита, а это вызывает переход воды из плазмы через поверхностную мембрану эритроцита. В результате объем эритроцитов в капиллярах большого круга увеличивается. При исследовании с помощью гематокрнта установлено, что эритроциты занимают 40% объема артериальной крови и 40,4% объема венозной крови. Из этого следует, что объем эритроцитов венозной крови больше, чем эритроцитом артериальной, что объясняется проникновением в них воды.
Одновременно с поступлением СО2 внутрь эритроцита и образованием в нем угольной кислоты происходит отдача кислорода оксигемоглобином и превращение его в редуцированный гемоглобин. Последний является значительно менее диссоциирующей кислотой, чем оксигемоглобин и угольная кислота. Поэтому при превращении оксигемоглобина в гемоглобин Н2СО3 вытесняет из гемоглобина ионы калия и, соединяясь с ними, образует калиевую соль бикарбоната.
Освобождающийся Н˙ ион угольной кислоты связывается гемоглобином. Так как редуцированный гемоглобин является малодиссоциированной кислотой, то при этом не происходит закисления крови и разница рН венозной и артериальной крови крайне невелика. Происходящую в эритроцитах тканевых капилляров реакцию можно представить следующим образом:
КНbO2 + Н2СO3= HHb + O2 + КНСO3
Из изложенного следует, что оксигемоглобин, превращаясь в гемоглобин и отдавая связанные им основания углекислоте, способствует образованию бикарбоната и транспорту в таком виде углекислоты. Кроме того, гкмоглобин образует химическое соединение с СО2 - карбогемоглобин. Наличие в крови соединения гемоглобина с углекислым газом было установлено путем следующего опыта. Если к цельной крови прибавить цианистый калий, который полностью инактивирует карбоангидразу, то оказывается, что эритроциты такой крови связывают больше СО2, чем плазма. Отсюда был сделан вывод, что связывание СО2 эритроцитами после инактивирования карбоангидразы объясняется наличием в эритроцитах соединения гемоглобина с СО2. В дальнейшем выяснилось, что СО2 присоединяется к аминной группе гемоглобина, образуя так называемую карбаминовую связь.
Реакция образования карбогемоглобина может идти в одну или другую сторону в зависимости от напряжения углекислого газа в крови. Хотя небольшая часть всего количества углекислого газа, которое может быть извлечено из крови, находится в соединении с гемоглобином (8-10%), однако роль этого соединения в транспорте углекислоты кровью достаточно велика. Примерно 25-30% углекислого газа, поглощаемого кровью в капиллярах большого круга, вступает в соединение с гемоглобином, образуя карбогемоглобин.
Отдача СО2 кровью в легочных капиллярах. Вследствие более низкого парциального давления СО2 в альвеолярном воздухе по сравнению с напряжением его в венозной крови углекислый газ переходит путем диффузии из крови легочных капилляров в альвеолярный воздух. Напряжение СО2в крови падает.
Одновременно с этим вследствие более высокого парциального давления кислорода в альвеолярном воздухе по сравнению с его напряжением в венозной крови кислород поступает из альвеолярного воздуха в кровь капилляров легких. Напряжение О2 в крови возрастает, и гемоглобин превращается в оксигемоглобин. Так как последний является кислотой, диссоциация которой значительно выше, чем гемоглобина угольной кислоты, то он вытесняет угольную кислоту из ее калиевой. Реакция идет следующим образом:
ННb + O2 + КНСO3= КНbO2+H2CO3
Освободившаяся из своей связи с основаниями угольная кислота расщепляется карбоангидразой на углекислый газ в воду. Значение карбоангидразы в отдаче углекислого газа в легких видно из следующих данных. Для того чтобы произошла реакция дегидратации Н2СО3 растворенной в воде, с образованием того количества углекислого газа, которое выходит из крови за время ее нахождения в капиллярах легких, требуется 300 секунд. Кровь же проходит через капилляры легких в течение 1-2 секунд, но за это время успевает произойти дегидратация угольной кислоты внутри эритроцита и диффузия образовавшегося СО2 сначала в плазму крови, а затем в альвеолярный воздух.
Так как в легочных капиллярах уменьшается в эритроцитах концентрация ионов НСО3, то эти ионы из плазмы начинают диффундировать в эритроциты, а ионы хлора диффундируют из эритроцитов в плазму. В связис тем что напряжение углекислого газа в крови легочных капилляров уменьшается, карбаминовая связь расщепляется и карбогемоглобин отдает углекислый газ.
Кривые диссоциации соединений угольной кислоты в крови. Как мы уже говорили, свыше 85% углекислого газа, которое может быть извлечено из крови подкислении ее, освобождается в результате расщепления бикарбонатов (калия в эритроцитах и натрия в плазме).
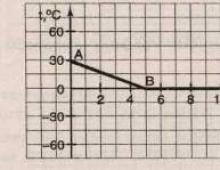
Связывание углекислого газа и отдача его кровью зависят от его парциального напряжения. Можно построить кривые диссоциации соединений углекислоты в крови, подобные кривым диссоциации оксигемоглобина. Для этого по оси ординат откладывают объемные проценты связанного кровью углекислого газа, а по оси абсцисс- парциальные напряжения углекислого газа. Нижняя кривая нарис. 58 показывает связывание углекислого газа артериальной кровью, гемоглобин которой почти полностью насыщен кислородом. Верхняя кривая показывает связывание кислого газа венозной кровью.
Различие в высоте этих кривых зависит от того, что артериальная кровь, богатая оксигемоглобином, обладает меньшей способностью связывать углекислый газ по сравнению с венозной кровью. Являясь более сильной кислотой, чем угольная кислота, оксигемоглобин отнимает основания у бикарбонатов и этим способствует освобождению угольной кислоты. В тканях оксигемоглобин, переходя в гемоглобин, отдает связанные с ним основания, увеличивая связывание кислого газа кровью.
Точка А на нижней кривой на рис. 58 соответствует напряжению кислоты, равному 40 мм рт. ст., т. е. тому напряжению, которое фактически имеется в артериальной крови. При таком напряжении связано 52 об.% СО2. Точка V на верхней кривой соответствует напряжению кислого газа 46 мм рт. ст., т. е. фактически имеющемуся в венозной крови. Как видно из кривой, при таком напряжении венозная кровь связывает 58 об.% углекислого газа. Линия AV, соединяющая верхнюю и нижнюю кривую, соответствует тем изменениям способности связывать углекислый газ, которые происходят при превращении артериальной крови в венозную или, наоборот, венозной крови в артериальную.
Венозная кровь благодаря тому, что содержащийся в ней гемоглобин переходит в оксигемоглобин, в капиллярах легких отдает около 6 об.% СО2. Если бы в легких гемоглобин не превращался в оксигемоглобин, то, как видно из кривой, венозная кровь при имеющемся в альвеолах парциальном давлении углекислого газа, равном 40 мм рт. ст.. связывала бы 54 об.% СО2, следовательно, отдала бы не 6, а только 4об.%. Равным образом, если бы артериальная кровь в капиллярах большого круга не отдавала своего кислорода, т. е. если бы гемоглобин ее оставался насыщенным кислородом, то эта артериальная кровь при парциальпом давлении углекислого газа, имеющемся в капиллярах тканей тела, смогла бы связат не 58 об.% СО2, а лишь 55 об.%.
234. Транспорт углекислоты кровью, значение карбоангидразы, взаимосвязь транспорта о2и со2.
Углекислый газ транспортируется следующими путями:
Растворенный в плазме крови - около 25 мл / л.
Связанный с гемоглобином (карбгемоглобин) - 45 мл / л.
В виде солей угольной кислоты - букарбонаты калия и натрия в плазме крови - 510 мл / л.
Таким образом, в состоянии покоя кровь транспортирует 580 мл углекислого газа в 1 л. Итак, основной формой транспорта СО2 является бикорбонаты плазмы, образующихся благодаря активному протеканию карбоангидразнои реакции.
В эритроцитах содержится фермент карбоангидраза (КГ), который катализирует взаимодействие углекислого газа с водой с образованием угольной кислоты, распадается с образованием бикарбонатного иона и протона. Бикарбонат внутри эритроцита взаимодействует с ионами калия, выделяемых из калиевой соли гемоглобина при восстановлении последнего. Так внутри эритроцита образуется бикарбонат калия. Но бикарбонатно ионы образуются в значительной концентрации и поэтому по градиенту концентрации (в обмен на ионы хлора) поступают в плазму крови. Так в плазме образуется бикарбонат натрия. Протон, образовавшегося при диссоциации угольной кислоты, реагирует с гемоглобином с образованием слабой кислоты ННb.
В капиллярах легких эти процессы идут в обратном направлении. С ионов водорода и бикарбонатных ионов образуется угольная кислота, которая быстро распадается на углекислый газ и воду. Углекислый газ удаляется наружу.
Итак, роль эритроцитов в транспорте углекислоты такова:
образование солей угольной кислоты;
образования карбгемоглобин.
Диффузия газов в тканях подчиняется общим законам (объем диффузии прямо пропорционален площади диффузии, градиента напряжения газов в крови и тканях). Площадь диффузии увеличивается, а толщина диффузного слоя уменьшается при увеличении количества функционирующих капилляров, что имеет место при повышении уровня функциональной активности тканей. В этих же условиях возрастает градиент напряжения газов за счет снижения в активно работающих органах Ро2 и повышения Рсо2 (газовый состав артериальной крови, как и альвеолярного воздуха остается неизменным!). Все эти изменения в активно работающих тканях способствуют увеличению объема диффузии О2 и СО2 в них. Потребление О2 (СО2) по спирограмму определяют по изменению (сдвигу) кривой вверх за единицу времени (1 минуту).
235. Иннервация дыхательных мышц.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга , иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровнеIII-IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположеныв передних рогах (III-XII) грудных сегментов спинного мозга.
236. Дыхательный центр. Современные представления о структуре и локализации. Автоматия дыхательного центра.
Информация о состоянии кислородно-углекислого баланса в организме поступает в дыхательный центр, который представляет нейронную организацию центральной нервной системы, определяющую функцию дыхания.
В анатомическом смысле дыхательный центр – это совокупность нейронов в локальной зоне центральной нервной системы, без которой дыхание становится невозможным.
Такой центр находится в ретикулярной формации продолговатого мозга в областидна IV желудочка .
Он состоит из двух отделов:
1) центр вдоха (инспираторный отдел);
2) центр выдоха (экспираторный отдел).
Нейроны бульбарного центра обладают автоматией и находятся в реципрокных взаимоотношениях между собой.
Несовершенность координации дыхательного акта центрами продолговатого мозга была доказана методом перерезок. Так после отделения продолговатого мозга от вышележащих отделов чередование вдохов и выдохов сохраняется, но длительность и глубина дыханий становится нерегулярной.
В физиологическом смысле дыхательный центр – это совокупность нейронов, расположенных на различных уровнях центральной нервной системы (от спинного мозга до коры головного мозга), которые обеспечивают координированное ритмическое дыхание, то есть делают функцию дыхания более совершенной.
В целом, регуляция активности дыхательного центра может быть представлена тремя уровнями:
1) на уровне спинного мозга располагаются центры диафрагмальных и межрёберных нервов, обусловливающие сокращение дыхательных мышц. Однако этот уровень регуляции дыхания не может обеспечить ритмическую смену фаз дыхательного цикла, так как большое количество афферентных импульсов от дыхательного аппарата непосредственно направляются в продолговатый мозг, то есть минуя спинной мозг.
2) на уровне продолговатого мозга и варолиевого моста находится основной дыхательный центр, который перерабатывает разнообразные афферентные импульсы, идущие от дыхательного аппарата, а также от основных сосудистых рефлексогенных зон. Этот уровень регуляции обеспечивает ритмическую смену фаз дыхания и активность спиномозговых мотонейронов, аксоны которых иннервируют дыхательную мускулатуру;
3) на уровне верхних отделов головного мозга , включая кору головного мозга, осуществляются адекватные приспособительные реакции системы дыхания к изменяющимся условиям окружающей среды.
Ритмические импульсы от дыхательного центра продолговатого мозга поступают по нисходящим двигательным путям к мотонейронам дыхательных мышц спинного мозга.
Мотонейроны диафрагмальных нервов находятся в передних рогах серого веществаIII - IV шейных сегментов .
Мотонейроны межрёберных нервов расположены в передних рогахгрудного отдела спинного мозга.
Отсюда возбуждение поступает к дыхательной мускулатуре (к диафрагме и межрёберным мышцам).
Мотонейроны спинного мозга
Бульбарный дыхательный центр
Мотонейроны спинного мозга получают от проприорецепторов мышц грудной клетки сигналы о степени их растяжения при вдохе.
Эти сигналы могут изменять число вовлечённых в активность мотонейронов и, таким образом, определяют особенности дыхания, осуществляя регуляцию дыхания на уровне спинного мозга
Бульбарный дыхательный центр получает афферентные импульсы от механорецепторов лёгких, дыхательных путей и дыхательных мышц, от хемо- и прессорецепторов сосудистых рефлексогенных зон.
Для нормальной деятельности бульбо-понтинного дыхательного центра необходима постоянная информация о состоянии внутренней среды организма и самих органов дыхания.
Нисходящие нервные влияния на дыхательный центр оказывают верхние отделы головного мозга , включая корковые нейроны. Так, эмоциональные возбуждения, охватывающие структуры,лимбико-ретикулярного комплекса и в первую очередьгипоталамическую область , распространяются в нисходящем направлении и вызывают изменение деятельности дыхательного центра.
Гипоталамус также оказывает влияния при изменениях во внешней среде, изменении метаболизма, а также как высший центр вегетативных регуляций.
Речь, относящаяся к высшим мозговым функциям коры человека, возможна на основе дыхательных движений, вызывающих прохождение воздуха через голосовой аппарат.
Поэтому во время речи к дыхательному центру приходят влияния, подстраивающие его деятельность для необходимых речевых реакций.
Одновременно дыхательный центр управляет тем объёмом лёгочной вентиляции, который необходим для поддержания дыхательного гомеостаза. Поэтому дыхание в условиях речи становится апериодическим.
На роль коры в регуляции дыхания указывает возможность произвольного контроля дыхания, когда человек может сознательно изменить дыхание: сделать его более глубоким или поверхностным, частым или редким, произвести задержку дыхания на определённое время.
Таким образом, на примере особенностей дыхательного центра наблюдаются общие принципы организации любых нервных центров, в частности:
1) принцип изоморфизма (принципиально однотипная структурная организация);
2) принцип иерархичности (многоуровневое расположение центрального представительства);
3) принцип субординации (соподчинение нервных центров, когда высшие центры модулируют работу низших и, чем выше уровень центра, тем более сложную регуляцию он обеспечивает).
Карбоангидраза
I
Карбоангидра́за (синоним: карбонатдегидратаза, карбонатгидролиаза)
фермент, катализирующий обратимую реакцию гидратации диоксида углерода: СО 2 + Н 2 О ⇔ Н 2 СО 3 ⇔ Н + + НСО 3 . Содержится в эритроцитах, клетках слизистой оболочки желудка, коре надпочечников, почках, в незначительных количествах - в ц.н.с., поджелудочной железе и других органах. Роль К. в организме связана с поддержанием кислотно-щелочного равновесия (Кислотно-щелочное равновесие),
транспортом СО 2 , образованием соляной кислоты слизистой оболочкой желудка.
К. в крови в норме довольно постоянна, но при некоторых патологических состояниях она резко меняется. Повышение активности К. в крови отмечается при анемиях различного генеза, нарушениях кровообращения II-III степени, некоторых заболеваниях легких (бронхоэктазах, пневмосклерозе), а также при беременности. Снижение активности этого фермента в крови происходит при ацидозе почечного генеза, гипертиреозе. При внутрисосудистом гемолизе К. появляется в моче, в то время как в норме она отсутствует. Контролировать активность К. в крови целесообразно во время оперативных вмешательств на и легких, т.к. она может служить показателем адаптивных возможностей организма, а также при терапии ингибиторами карбоангидразы - гипотиазидом, диакарбом. Для определения активности К. применяют радиологические, иммуноэлектрофоретические, колориметрические и титриметрические методы. Определение производят в цельной крови, взятой с гепарином, или в гемолизированных эритроцитах. Для клинических целей наиболее приемлемы колориметрические методы определения активности К. (например, модификации метода Бринкмана), основанные на установлении времени, необходимого для сдвига рН инкубационной смеси с 9,0 до 6,3 в результате гидратации СО 2 . Воду, насыщенную углекислотой, смешивают с индикаторно-буферным раствором и определенным количеством сыворотки крови (0,02 мл
) или взвеси гемолизированных эритроцитов. В качестве индикатора используют феноловый красный. По мере диссоциации молекул угольной кислоты все новые молекулы СО 2 подвергаются ферментативной гидратации. Для получения сравнимых результатов должна протекать всегда при одинаковой температуре, наиболее удобно поддерживать температуру тающего льда - 0°. Время контрольной реакции (спонтанной реакции гидратации СО 2) в норме составляет 110-125 с
. В норме при определении этим методом активность К. в среднем равна 2-2,5 условным единицам, а в пересчете на 1 млн. эритроцитов 0,458 ± 0,006 условным единицам (за единицу активности К. принимают увеличение скорости катализируемой реакции в 2 раза). Библиогр.:
Клиническая оценка лабораторных тестов, под ред. Н.У. Тица, . с англ., с. 196, М., 1986.
1. Малая медицинская энциклопедия. - М.: Медицинская энциклопедия. 1991-96 гг. 2. Первая медицинская помощь. - М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. - М.: Советская энциклопедия. - 1982-1984 гг .
Синонимы :Смотреть что такое "Карбоангидраза" в других словарях:
Карбоангидраза … Орфографический словарь-справочник
Фермент, катализирующий обратимую реакцию образования угольной кислоты из диоксида углерода и воды. Ингибиторы карбоангидразы применяют в медицине для лечения некоторых сердечно сосудистых и др. заболеваний … Большой Энциклопедический словарь
Угольная ангидраза, карбонат гидролиаза, фермент класса лиаз, катализирует обратимую реакцию гидратации двуокиси углерода. Обнаружена у животных, человека, растений, бактерий. Содержит в качестве кофактора атом Zn. Мол. м. 28 000 30 000.… … Биологический энциклопедический словарь
Сущ., кол во синонимов: 1 фермент (253) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
карбоангидраза - Металлофермент (содержит в активном центре ионы цинка), катализирущий обратимую реакцию гидратирования углекислоты; дефицит К. является причиной мраморной болезни у человека. [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь… … Справочник технического переводчика
Фермент, катализирующий обратимую реакцию образования угольной кислоты из диоксида углерода и воды. Ингибиторы карбоангидразы применяют в медицине для лечения некоторых сердечно сосудистых и других заболеваний. * * * КАРБОАНГИДРАЗА КАРБОАНГИДРАЗА … Энциклопедический словарь - угольная ангидраза, карбонат гидролиаза, фермент класса лиаз (См. Лиазы), катализирующий обратимое образование угольной кислоты из двуокиси углерода и воды: CO2 + H2O ↔ H2CO3. К. металлопротеид, содержащий Zn; молекулярная масса около 30… … Большая советская энциклопедия
Фермент, катализирующий обратимую реакцию образования угольной кислоты из диоксида углерода и воды. Ингибиторы К. применяют в медицине для лечения нек рых сердечно сосудистых и др. заболеваний … Естествознание. Энциклопедический словарь
Первые школьные уроки об устройстве человеческого организма знакомят с главными «обитателями крови: красные клетки – эритроциты (Er, RBC), определяющие цвет за счет , в них содержащегося, и белые (лейкоциты), присутствие которых на глаз не видно, поскольку на окраску они не влияют.
Эритроциты человека, в отличие от животных, не имеют ядра, но прежде чем потерять его, они должны пройти путь от клетки-эритробласта, где только начинается синтез гемоглобина, достигнуть последней ядерной стадии – , накапливающего гемоглобин, и превратиться в зрелую безъядерную клетку, основным компонентом которой является красный кровяной пигмент.

Чего только люди не делали с эритроцитами, изучая их свойства: и вокруг земного шара пытались их обернуть (получилось 4 раза), и в монетные столбики укладывать (52 тысячи километров), и площадь эритроцитов сопоставлять с площадью поверхности тела человека (эритроциты превзошли все ожидания, их площадь оказалась выше в 1,5 тысячи раз).
Эти уникальные клетки…
Еще одна важная особенность эритроцитов заключается в их двояковогнутой форме, но если бы они были шарообразными, то общая площадь их поверхности была бы меньше на 20% настоящей. Однако способности эритроцитов заключаются не только в величине их общей площади. Благодаря двояковогнутой дисковидной форме:
- Эритроциты способны переносить больше кислорода и углекислого газа;
- Проявлять пластичность и свободно проходить через узкие отверстия и изогнутые капиллярные сосуды, то есть, для молодых полноценных клеток в кровяном русле практически нет препятствий. Способность проникать в самые отдаленные уголки организма теряется с возрастом эритроцитов, а также при их патологических состояниях, когда изменяется их форма и размер. Например, сфероциты, серповидные, гири и груши (пойкилоцитоз), не обладают такой высокой пластичностью, не могут пролезать в узкие капилляры макроциты, а тем более, мегалоциты (анизоцитоз), поэтому и задачи свои измененные клетки выполняют не столь безупречно.

Химический состав Er представлен в большей степени водой (60%) и сухим остатком (40%), в котором 90 – 95% занимает красный пигмент крови – , а остальные 5 – 10% распределены между липидами (холестерин, лецитин, кефалин), белками, углеводами, солями (калий, натрий, медь, железо, цинк) и, конечно, ферментами (карбоангидраза, холинэстераза, гликолитические и пр.).
Клеточные структуры, которые мы привыкли отмечать в других клетках (ядро, хромосомы, вакуоли), у Er отсутствуют за ненадобностью. Живут эритроциты до 3 – 3,5 месяцев, затем состариваются и с помощью эритропоэтических факторов, которые выделяются при разрушении клетки, подают команду, что их пора заменить новыми – молодыми и здоровыми.
Начало свое эритроцит берет от предшественников, которые, в свою очередь, происходят от стволовой клетки. Воспроизводятся красные кровяные тельца, если в организме все нормально, в костном мозге плоских костей (череп, позвоночник, грудина, ребра, тазовые кости). В случаях, когда по каким-либо причинам костный мозг не может их производить (поражение опухолью), эритроциты «вспоминают», что во внутриутробном развитии этим занимались другие органы (печень, вилочковая железа, селезенка) и заставляют организм начать эритропоэз в забытых местах.
Сколько их должно быть в норме?
Общее количество эритроцитов, содержащееся в организме в целом, и концентрация красных клеток, курсирующих по кровяному руслу – понятия разные. В общее число входят клетки, которые еще пока не покинули костный мозг, ушли в депо на случай непредвиденных обстоятельств или пустились в плавание для выполнения своих непосредственных обязанностей. Совокупность всех трех популяций эритроцитов носит название – эритрон . В эритроне содержится от 25 х 10 12 /л (Тера/литр) до 30 х 10 12 /л красных кровяных клеток.

Норма эритроцитов в крови взрослых людей отличается по половому признаку, а у детей в зависимости от возраста. Таким образом:
- Норма у женщин колеблется в пределах 3,8 – 4,5 х 10 12 /л, соответственно, гемоглобина у них тоже меньше;
- Что для женщины является нормальным показателем, то у мужчин называется анемией легкой степени, поскольку нижняя и верхняя граница нормы эритроцитов у них заметно выше: 4,4 х 5,0 х 10 12 /л (то же самое касается и гемоглобина);
- У детей до года концентрация эритроцитов постоянно меняется, поэтому для каждого месяца (у новорожденных – каждого дня) существует своя норма. И если вдруг в анализе крови повышены эритроциты у ребенка двух недель отроду до 6,6 х 10 12 /л, то это нельзя расценивать как патологию, просто у новорожденных такая норма (4,0 – 6,6 х 10 12 /л).
- Некоторые колебания наблюдаются и после года жизни, но нормальные значения не особо отличаются от таковых у взрослых. У подростков 12 -13 лет содержание гемоглобина в эритроцитах и уровень самих эритроцитов соответствует норме взрослых людей.
Повышенное содержание эритроцитов в крови называется эритроцитозом , который бывает абсолютным (истинным) и перераспределительным. Перераспределительный эритроцитоз патологией не является и возникает, когда эритроциты в крови повышены при определенных обстоятельствах:
- Пребывание в горной местности;
- Активный физический труд и спорт;
- Психоэмоциональное возбуждение;
- Дегидратация (потеря организмом жидкости при диарее, рвоте и т. д.).
Высокие показатели содержания эритроцитов в крови являются признаком патологии и истинного эритроцитоза, если они стали результатом усиленного образования красных кровяных телец, вызванного неограниченной пролиферацией (размножением) клетки-предшественницы и ее дифференцировки в зрелые формы эритроцитов ().
Снижение концентрации красных клеток крови называют эритропенией . Она наблюдается при кровопотере, угнетении эритропоэза, распаде эритроцитов () под действием неблагоприятных факторов. Низкие эритроциты в крови и пониженное содержание Hb в эритроцитах является признаком .
О чем говорит аббревиатура?
Современные гематологические анализаторы, помимо гемоглобина (HGB), пониженного или повышенного содержания эритроцитов в крови (RBC), (HCT) и других привычных анализов, могут рассчитывать и другие показатели, которые обозначаются латинской аббревиатурой и бывают совсем не понятны читателю:

Кроме всех перечисленных достоинств эритроцитов, хочется отметить еще одно:
Эритроциты считают зеркалом, отражающим состояние многих органов. Своеобразным индикатором, способным «почувствовать» неполадки или позволяющим следить за течением патологического процесса, является .
Большому кораблю – большое плавание
Почему красные кровяные клетки так важны для диагностики многих патологических состояний? Их особая роль вытекает и формируется в силу уникальных возможностей, а чтобы читатель мог себе представить истинную значимость эритроцитов, попробуем перечислить их обязанности в организме.

Поистине, функциональные задачи красных кровяных клеток широки и многообразны:
- Они осуществляют транспортировку кислорода к тканям (с участием гемоглобина).
- Переносят углекислый газ (с участием, помимо гемоглобина, фермента карбоангидразы и ионообменника Cl- /HCO 3).
- Выполняют защитную функцию, так как способны адсорбировать вредные вещества и переносить на своей поверхности антитела (иммуноглобулины), компоненты комплементарной системы, образованные иммунные комплексы (Ат-Аг), а также синтезировать антибактериальное вещество, называемое эритрином .
- Участвуют в обмене и регуляции водно-солевого равновесия.
- Обеспечивают питание тканей (эритроциты адсорбируют и переносят аминокислоты).
- Участвуют в поддержании информационных связей в организме за счет переноса макромолекул, которые эти связи обеспечивают (креаторная функция).
- Содержат тромбопластин, который выходит из клетки при разрушении эритроцитов, что является сигналом для системы свертывания начать гиперкоагуляцию и образование . Кроме тромбопластина, эритроциты несут гепарин, препятствующий тромбообразованию. Таким образом, активное участие эритроцитов в процессе свертывания крови – очевидно.
- Красные клетки крови способны подавлять высокую иммунореактивность (выполняют роль супрессоров), что может быть использовано в лечении различных опухолевых и аутоиммунных заболеваний.
- Участвуют в регуляции производства новых клеток (эритропоэз) путем освобождения из разрушенных старых эритроцитов эритропоэтических факторов.
Разрушаются красные кровяные тельца преимущественно в печени и селезенке с образованием продуктов распада ( , железо). Кстати, если рассматривать каждую клетку по отдельности, то она будет не такой уж и красной, скорее, желтовато – красной. Скапливаясь в огромные миллионные массы, они, благодаря гемоглобину, в них находящемуся, становятся такими, как мы привыкли их видеть – насыщенно-красного цвета.
Видео: урок по эритроцитам и функциям крови